Птица с руками и птица-кенгуру
Удивительная птица Гоацин живет в тропических лесах бассейна Амазонки. Это небольшая птичка размером с цыпленка имеет очень тонкую шейку. Но самое необыкновенное в этой птичке то, что вместо привычных двух лап у нее имеются дополнительные, которые очень напоминают руки.
Именно так видоизменились крылья гоацина в процессе эволюции. Ученые орнитологи выявили сходство гоацина с древней птицей археоптриксом. С помощью своих «рук» гоацин легко и цепко хватается за ветки деревьев и может висеть на них продолжительное время. Еще одним резким отличием от своих пернатых сородичей является то, что гоацин не поет, а как лягушка квакает. Мясо гоацина не пригодно для еды, а все хищников он отпугивает резким неприятным запахом.
Птица-кенгуру
Редкая водоплавающая птица встречается в лесах Южной и Центральной Америки. Ее называют Пикапаре или файнфут. Название файнфут имеет английские корни и означает роскошные ноги. Так его называют из-за желтых ног с красивыми черными полосами.
Но самой главной особенностью пикапаре является то, что это единственная птица, которая переносит своих птенчиков в удиветельных мешочках под крыльями. Это очень похоже на то как кенгуру носит своих детенышей в сумке на животе, но в отличие от австралийского «собрата» птенцов переносит самец, а не самка. Мешочки расположены под крыльями и выложены перьями, поэтому даже на дальних перелетах птенцам удобно сидеть в этих мешочках.
В перьевых карманах содержится достаточное количество воздуха, чтобы
отец мог нырять за добычей вместе с птенцами. Едой пикапаре служит рыба
и мелкие организмы, обитающие в водоемах и речными растениями.
Птицы — Википедия
Пти́цы (лат. Aves) — группа теплокровных яйцекладущих позвоночных животных, традиционно рассматриваемая в ранге отдельного класса[К 1]. Представляют собой хорошо обособленную на фоне остальных современных животных группу, одним из наиболее характерных признаков представителей которой является покров из перьев, предохраняющий тело от неблагоприятных изменений температуры и играющий важную роль при полёте[1]. Способность к полёту является главнейшей особенностью птиц (отсутствие её у некоторых видов — вторичное явление)[2]. Передние конечности приобрели форму крыльев. Птицы обладают особым строением органов дыхания и пищеварения, что тесно связано с их способностью летать. Ещё одним отличительным признаком является наличие клюва[1][2].
Многие виды птиц совершают регулярные длительные миграции из одного региона Земли в другой. Ещё большее количество ведёт кочевой образ жизни, постоянно перемещаясь на небольшие расстояния в пределах своих ареалов. Птицы являются социальными животными, которые общаются между собой при помощи визуальных и звуковых сигналов, и выполняют в ряде случаев общественные действия: коммунальное гнездование, совместная охота или защита от хищников
Птицы населяют все экосистемы земного шара, включая внутренние части Антарктиды[1]. Согласно данным Международного союза орнитологов, на декабрь 2019 года науке известно &&&&&&&&&&010758.&&&&&010 758 видов ныне живущих птиц (&&&&&&&&&&020034.&&&&&020 034 подвидов), а также 158 видов, вымерших в современное историческое время [3]. Это делает птиц наиболее разнообразной группой надкласса четвероногих. На территории России отмечено 789 видов, включая 657 гнездящихся видов; 125 видов, гнездование которых не показано, и 7 вымерших видов[4].
Будучи наиболее многочисленной и широко распространенной группой высших позвоночных, а также вследствие особенностей своей биологии, птицы играют важную роль в природе и в жизни человека. Многие виды имеют важное экономическое значение. Птицеводство является одной из основных отраслей сельского хозяйства, производящей для человека мясо, яйца и жир, а также перья. Многие виды птиц содержатся человеком в качестве домашних животных. Вследствие деятельности человека многие птицы получили своё дальнейшее развитие, некоторые стали синантропами, но в то же время около 1200 видов в той или иной степени подвержены риску вымирания и охраняются национальными и международными законами
Наука, занимающаяся изучением птиц, называется орнитологией[6].
Русское слово «птица» восходит к праслав. *ръtica, которое является суффиксальным производным от *ръtа, родственного латыш. putns «птица», лит. putýtis «пташка», лат. putus «дитя», putila «птенец», оскск. рuclо- «дитя», др.-инд. पुत्रः (putráḥ IAST) «дитя, сын», др.-инд. पोतः (рótаḥ IAST) «детёныш животного», и восходящего в свою очередь к пра-и.е. *pōu-/*pəu-/*pu-/*pū- «маленький»[7][8].
Латинское слово avis восходит к пра-и.е. *h2ewis (в традиционной реконструкции *awis) «птица», от которого происходят также др.-инд. विः (viḥ IAST) «птица», др.-греч. ἀετός «орёл», арм. հավ (hav) «курица», валл. hwyad «утка»[9][10].
Размеры и масса тела птиц различны. Масса крупных летающих птиц обычно не превышает 12—16 кг [6][11] (лебеди, грифы, дрофы)[6] при размахе крыльев до 3 м (пеликаны, альбатросы)[6][12]. Самые мелкие из птиц — некоторые виды колибри — имеют максимальную массу тела 3—7 г[13]. Утрата способности к полёту часто приводит к увеличению размеров[6]: масса крупных пингвинов достигает 40 кг, казуаров и африканских страусов — 80—150 кг[14]. Некоторые вымершие нелетающие птицы достигали массы 230—400 кг (эпиорнисовые, моа)[15].
Самой маленькой из ныне живущих птиц считается колибри-пчёлка — длина тела не превышает 5,7 см[16]. Самой крупной современной птицей является африканский страус с высотой до 2,7 м и массой до 156 кг[12][15]. Таким образом, масса самой маленькой птицы в 66 тысяч раз меньше массы самой крупной из ныне живущих[14].
Андский кондор принадлежит к числу наиболее крупных летающих птиц — масса тела достигает до 11,4 кг при размахе крыльев до 3,1 м
Странствующий альбатрос, достигая длины тела до 117 см, обладает самым большим среди ныне живущих птиц размахом крыльев — до 3,25 м[12][18].
-

Колибри-пчёлка
-
Африканский страус
-
Андский кондор
Странствующий альбатрос
-
Сравнительный размер Pelagornis sandersi с современными андским кондором и странствующим альбатросом




Наибольшим размахом крыльев, который по различным оценкам достигал 6,1—7,4 м, обладал ископаемый вид Pelagornis sandersi из олигоцена[19]. До его описания в 2014 году крупнейшей летающей птицей считался ископаемый аргентавис с размахом крыльев около 7 м, известный из миоцена Аргентины[20].
Происхождение[править | править код]
 Археоптерикс. Берлинский экземпляр
Археоптерикс. Берлинский экземплярПроисхождение птиц долгое время оставалось предметом оживлённых дискуссий[12]. За обозримый период времени было выдвинуто несколько научных версий происхождения и родственных связей птиц и возникновения у них полёта, причём в течение более сотни лет они были чисто гипотетическими
Впервые теория эволюции птиц от рептилий возникла после открытия в 1860 году в Германии ископаемых остатков археоптерикса — животного, жившего около 150 млн лет назад в верхней юре. Он обладал характеристиками типичного пресмыкающегося — особым строением таза и рёбер, зубами, когтистыми лапами и длинным, как у ящерицы, хвостом. При этом окаменелости имели хорошо сохранившиеся отпечатки маховых перьев, аналогичные таковым у современных птиц. На протяжении многих десятилетий история развития птиц рассматривалась как эволюция группы, развивавшейся от археоптерикса.
Именно на его изучении и основывались все первые гипотезы и теории о происхождении и родственных связях птиц: теория древесного («с деревьев вниз», Marsh, 1877)[22] и теория бегательного («с земли вверх», Williston, 1879) [23] возникновения полёта у птиц. Соответственно этим представлениям трактовалось и происхождение самих птиц — от триасовых текодонтов (архозавроморф) при древесной теории или от юрских бегающих тероподных динозавров при наземной теории[24].
В настоящее время археоптерикса уже не рассматривают в качестве общего предка всех современных птиц. Однако он, вероятно, имеет близкое родство с их настоящим предком[25]. Точное положение археоптерикса в эволюционном дереве определить трудно. Согласно кладистическому анализу китайских палеонтологов Xu et al. 2011 года, археоптерикс может представлять собой (вместе с видом Xiaotingia zhengi) параллельную тупиковую ветвь на общем стволе динозавров[26]. Однако более тщательный филогенетический анализ не подтвердил размещение археоптерикса среди дейнонихозавров, и поэтому его продолжают считать древнейшей и наиболее примитивной птицей (в составе группы Avialae)[27]
Динозавры и птицы[править | править код]

Гипотеза о происхождении птиц от динозавров-тероподов впервые была выдвинута в 1868 году Томасом Гексли. Она основывалась на сравнении строения археоптерикса и маленького хищного динозавра рода Compsognathus.
С течением времени, и особенно в последние десятилетия, в связи с открытием многих видов древних птиц и оперённых динозавров-теропод большинство исследователей[24] стали склоняться к теории, согласно которой птицы произошли от динозавров-тероподов из группы манирапторов[29].
 Реконструкция летающего хищного динозавра чанъюйраптора, покрытого перьями
Реконструкция летающего хищного динозавра чанъюйраптора, покрытого перьямиПо мере обнаружения всё большего количество ископаемых остатков нелетающих теропод, так или иначе имеющих родство с птицами, чёткая граница между «птицами» и «не-птицами» становится размытой, и провести различие между данными группами всё сложнее[27]. Если ранее одним из определяющих признаков птиц считалось наличие перьевого покрова, то ряд открытий конца XX — начала XXI веков в провинции Ляонин на северо-востоке Китая показывает, что многие мелкие тероподы, например синозавроптерикс[30] и синорнитозавр[31], имели перья[32]. Самым ранним из известных «перьев», вероятно, является окаменелость, описанная под названием Praeornis sharovi[33][34][35]. Эволюционное происхождение перьев можно проследить начиная с таких крупных тероподов, как карнотавр, покрытого многочисленными бугорками[36], похожими на фолликулы. Следующий этап виден у синозавроптерикса и Dilong paradoxus, покрытых волокнистым пухом[36]. Наконец, у каудиптерикса, протархеоптерикса, синорнитозавра, микрораптора и Changyuraptor yangi уже имелись настоящие перья[36].
| Кладограмма филогении птиц согласно Godefroit et al., 2013[37] |
Птиц роднит с тероподами множество общих черт в строении скелета[12]. В последние годы среди палеонтологов достигнут консенсус, что ближайшими родственниками современных птиц являются дейнонихозавры — инфраотряд, включающий в себя в том числе семейства дромеозавров и троодонтид. Эти три группы были объединены в единую, получившую название Paraves.
Центральное место среди дромеозавров занимает «четырёхкрылый» микрораптор, который был способен летать либо планировать. Тот факт, что дейнонихозавры были в большинстве своём маленького размера, дал основание предполагать, что предок птиц вёл древесный образ жизни и был способен к планированию[38][39].
Среди возможных ближайших родственников птиц также рассматривается авимим — мелкий хищный динозавр из мелового периода[40].
Альтернативные версии происхождения[править | править код]
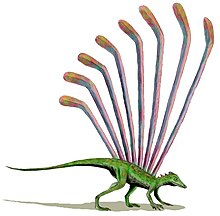
Наряду с теорией происхождения от динозавров существует гипотеза, согласно которой птицы могли эволюционировать от более ранних архозавроморфов[21] независимо от линии развития собственно динозавров, в поздней перми или раннем триасе[41]. Раньше в качестве такого предка рассматривались мелкие и более примитивные псевдозухии из группы, традиционно называемой «текодонтами» (сейчас это таксономическое название устарело), теперь возможными предками считают преимущественно орнитозухид[21]. Впервые данная теория была высказана в 1913—1914 годах Робертом Брумом и Фридрихом Хюне, позднее защищалась Герхардом Хейельманом и оставалась наиболее популярной вплоть до конца двадцатого столетия. Тем не менее, некоторые учёные, среди которых американские палеонтологи Ларри Мартин и Алан Федучия[en], продолжают настаивать, что птицы произошли от более ранних архозавров типа лонгисквамы[41]. Они утверждают, что сходство между птицами и манирапторами является результатом конвергентной эволюции. В конце 1990-х годов Федучия и Мартин выдвинули модифицированную версию своей гипотезы, согласно которой манирапторы являются нелетающими потомками ранних птиц[42]. Данная гипотеза оспаривается большинством палеонтологов[43]. В июле 2014 года в продолжение своей гипотезы Алан Федучия в соавторстве опубликовал исследование, согласно которому Scansoriopteryx рассматривался ими как древняя птица — потомок архозавров. По мнению авторов публикации, наличие у него архаичных черт и анатомических особенностей, характерных для птиц, и перьев на конечностях свидетельствуют о возникновении полёта «сверху вниз», при котором архозавры обзавелись перьями. Ими также утверждается, что Scansoriopteryx и все птицы произошли от архозавров, таких как склеромохлус[44]. Филогенетические исследования никогда не подтверждали данную альтернативную интерпретацию, и прежние аргументы авторов публикации ранее уже неоднократно были подвергнуты критике как ненаучные[43].
Ещё одну версию происхождения птиц отстаивал российский палеонтолог Евгений Курочкин. По его мнению, предком настоящих птиц мог быть протоавис, который на 75 млн лет древнее археоптерикса[21][45]. Критики данной теории утверждают, что ископаемые остатки протоависа плохо сохранились и могут быть химерой[46].
В 1977 году палеонтолог Алик Уокер после изучения скелета триасового южноафриканского текодонта-крокодиломорфа сфенозуха предложил гипотезу о происхождении птиц от ранних, позднетриасовых крокодиломорфов. Она базировалась на большом количестве общих черт строения черепа ранних крокодиломорфов и птиц[47]. Аргументы Уокера неоднократно критиковались, и сам автор впоследствии признал гипотезу несостоятельной, поскольку сходство птиц и крокодиломорфов ограничивается типичными признаками архозавров.
Помимо описанных выше основных теорий происхождения птиц, были выдвинуты и многие другие. Однако все эти гипотезы не принимаются сегодня всерьёз. К таковым относятся теория Фогта о родстве птиц и ящериц (1879), теория о связи птиц с птерозаврами (Оуэн, 1875; Сили, 1881), теория о происхождении птиц от эозухий (Г. П. Дементьев, 1940)[48], теория о родстве птиц и птицетазовых динозавров (Galton, 1970)[49].
Ранняя эволюция птиц[править | править код]
Конфуциусорнис — примитивная ископаемая птица из раннего мелового периода. КитайНаиболее ранние представители клады Avialae — птицеподобные динозавры Anchiornis huxleyi, Xiaotingia zhengi, Aurornis xui, известные по окаменелостям из геологической формации Тяоцзишань в Китае, датируемой концом юрского периода (около 160 млн лет назад)[37]. Многие из этих ранних форм обладали необычными анатомическими особенностями, которые в дальнейшем были утеряны птицами в ходе эволюции[37].
Эволюционное разделение на множество разнообразных форм началось в меловом периоде[51]. Многие из этих форм, в том числе и предки современных веерохвостых птиц, на первоначальном этапе сохраняли примитивные признаки ящеров — крылья с когтями и зубы, которые впоследствии были редуцированы. Джехолорнис и другие древнейшие птицы обладали длинным хвостовым отделом позвоночника[51]. У представителей других, более развитых групп, например, клады пигостилевых, 4—6 задних хвостовых позвонков были слиты воедино, образовав пигостиль.
Первой широко распространённой и разнообразной группой короткохвостых птиц стали энанциорнисовые, или «противоположные птицы», названные так потому, что строение их плечевых костей было противоположно строению таковых у современных птиц[52]. Примитивные энанциорнисовые не связаны общим происхождением с более прогрессивными веерохвостыми птицами и представляют отдельную ветвь эволюции. Среди внешне похожих на современных птиц и развивавшихся вместе с ними в меловом периоде энанциорнисов были представители, занявшие различные экологические ниши: от обитателей песчаных морских побережий и охотников за рыбой до ведущих древесный образ жизни и питающихся семенами[51]. Более развитые ихтиорнисы, несколько напоминающие современных чаек, обитали на морских просторах и также питались рыбой[53]. Морские птицы из отряда гесперорнисообразных из мезозоя настолько приспособились к охоте за рыбой, что в ходе эволюции утратили способность к полёту и полностью перешли к водному образу жизни. Несмотря на такую узкую специализацию, они являются одними из родственников современных птиц[51].
Эволюция современных птиц[править | править код]
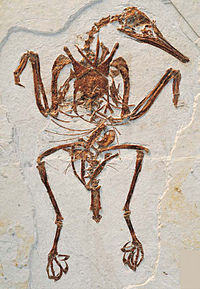
Обнаруженные на острове Вега у берегов Западной Антарктиды ископаемые остатки древней птицы вегавис[54] показали, что по крайне мере одна эволюционная линия веерохвостых птиц получила развитие в начале мелового периода[54][55] и впоследствии разделилась на две основные формы, известные сегодня как подклассы бескилевых и новонёбных птиц. Бескилевые представляют относительно небольшую группу, к которой относят страусообразных и скрытохвостых. Следующей группой, разошедшейся с остальными новонёбными птицами, стали представители надотряда Galloanserae, в состав которого включают гусеобразных и курообразных. Период, когда началось данное расхождение, является предметом научных споров — большинство специалистов считают, что веерохвостые получили своё развитие в меловом периоде, а ветвь Galloanserae отделилась от остальных новонёбных до начала мел-палеогенового вымирания. Тем не менее, у этой версии имеются и противники, полагающие, что расхождение могло произойти уже после (и благодаря) гибели большей части животного мира[56]. Разногласие отчасти вызвано различными методами исследований: молекулярная генетика указывает на основное увеличение разнообразия в середине мелового периода, а анализ окаменелостей свидетельствует о начале палеогена[56][57]. Последнее не удивительно, поскольку в первом случае речь идёт о времени отделения последнего общего предка данной группы от базальной, что произошло заведомо раньше появления остатков специализированных разновидностей в палеонтологической летописи, поскольку в неё попадают только весьма распространённые в данный момент времени организмы. Более того, на момент разделения предки различных групп птиц могли быть практически неотличимы друг от друга по морфологии — всё различие между ними состояло в обусловленном генофондом эволюционном потенциале (так же, как и у различных групп примитивных плацентарных млекопитающих, которые на момент первичной радиации в палеогене были почти неотличимы друг от друга, но впоследствии дали начало совершенно различным отрядам).

Древнейшей известной науке веерохвостой (настоящей) птицей является Archaeornithura meemannae, жившая около 130,7 млн лет назад на территории современного Китая[58][59].
Примерно 60—40 млн лет назад произошла интенсивная адаптивная радиация веерохвостых птиц, возникло большинство современных отрядов. Окаменелости птиц этого времени малочисленны и не дают представления о темпах и характере эволюции. Птицы, известные из кайнозоя, вероятно, имеют общего предка выше уровня зубастых Hesperornithiformes и
Пикайя — Википедия
| † Пикайя | |||||||
|---|---|---|---|---|---|---|---|
 | |||||||
промежуточные ранги
| |||||||
Pikaia gracilens Walcott, 1911 | |||||||
Пикайя[источник не указан 1123 дня] (лат. Pikaia gracilens) — небольшое примитивное хордовое животное. Ископаемые остатки пикайи встречаются в среднекембрийских отложениях возрастом 530 млн лет в сланцах Бёрджес (Канада).
Окаменелость тела пикайи Размеры тела пикайи в сравнении с другими представителями фауны сланцев Бёрджесс
Размеры тела пикайи в сравнении с другими представителями фауны сланцев БёрджессНаучное родовое название Pikaia дано по названию горы Pika Peak (англ.)русск.[1], расположенной в Канаде на границе провинции Альберта и Британской Колумбии. Название горы может быть переведено как Пищуховая вершина.
В 1909 году американский палеонтолог Ч. Д. Уолкотт обнаружил в среднекембрийских отложениях Британской Колумбии (Канада) в сланцах Бёрджесс многочисленные остатки ископаемых организмов. Многие из них принадлежали мягкотелым бесскелетным формам. В ходе последовавших за этим открытием раскопок было собрано большое количество палеонтологического материала. Среди найденных образцов были и «отпечатки» пикайи, фактически представлявшие собой тонкие углеродистые и алюмосиликатные плёнки на поверхности горной породы[2][3].
В 1911 году Ч. Д. Уолкотт опубликовал научное описание пикайи. Это животное было выделено им в самостоятельное семейство Pikaidae с единственным родом и видом и отнесено к многощетинковым червям — полихетам[1].
В конце 1970-х годов британский палеонтолог С. Конвей Моррис обнаружил у пикайи образование, напоминающее хорду, а также мускульные сегменты — миотомы, характерные для хордовых, поэтому большинство специалистов стало относить пикайю к хордовым[3]. В 2012 году С. Конвей Моррис совместно с канадским палеонтологом Ж.-Б. Кароном опубликовал новые данные по анатомии пикайи. В результате изучения всех обнаруженных к тому времени 114 образцов эти специалисты предложили новую интерпретацию строения тела данного животного[2][3].
Внешний вид[править | править код]
Длина тела взрослых особей достигала от 1,5 до 6 см, в среднем она равнялась 4 см. Высота тела составляла от 7 до 16 % его длины, в среднем около 11 %[2].
Тело пикайи удлинённой формы, округлого сечения в передней части и сжатое с боков в остальных частях. Вдоль брюшной стороны тянулся узкий гребень, напоминающий плавник. Ещё один гребень, хотя и не столь высокий, располагался на спинной стороне[2][3].
Голова была очень маленькая, подразделённая на две округлые лопасти, на которых имелась пара щупалец. Есть основания полагать, что эти щупальца были упругими, поскольку после смерти животного они обычно оставались прямыми, судя по сохранившимся отпечаткам. Скорее всего, они выполняли функцию органов обоняния и осязания. У основания головы на её нижней стороне размещалось ротовое отверстие. Никаких органов, похожих на глаза, не обнаружено[2][3].
Позади головы по обеим сторонам глотки имелось 9 пар ветвистых придатков, которые трактуются исследователями как наружные жабры. Вблизи оснований этих придатков иногда видны небольшие округлые пятна, возможно, соответствующие жаберным щелям[2][3].
Анатомия[править | править код]
Мускулатура разделена на множество поперечных сегментов — миотомов, которых насчитывается до сотни. Миомеры имеют сигмоидные очертания. Границы между миотомами хорошо заметны на ископаемых остатках, поскольку в соответствующих местах отпечатков повышено содержание углерода. По всей видимости, миотомы были разделены прослойками плотной соединительной ткани, которая разлагалась медленнее, чем мускулатура животного[2][3]. Сегментация распространяется и на брюшной плавник[2].
Внутри туловища на спинной стороне расположен плотный продольный тяж, который сохраняет объёмную, выпуклую форму даже у ископаемых остатков. Он начинается позади глотки и тянется до самого конца туловища. Раньше он рассматривался исследователями как хорда. Однако, его толщина слишком велика для хорды. Кроме того, он расположен слишком высоко, не оставляя места для нервной трубки, которая должна размещаться выше, чем хорда. Прежняя интерпретация была пересмотрена в 2012 году С. Конвеем Моррисом и Ж.-Б. Кароном. В их работе этот тяж именуется спинным органом. Происхождение и функции данного органа пока остаются неясными[2][3].
Хорда и нервная трубка, по интерпретации С. Конвея Морриса и Ж.-Б. Карона, располагаются ниже спинного органа[2][3].
Пищеварительная система открывается ротовым отверстием у основания головы. В передней части кишечника имеется расширение — глотка. Анальное отверстие, согласно реконструкции С. Конвея Морриса и Ж.-Б. Карона, расположено на заднем конце тела[2].
Ещё один тонкий продольный тяж расположен на брюшной стороне тела. С. Конвеем Моррисом и Ж.-Б. Кароном он интерпретируется как кровеносный сосуд. На некоторых экземплярах подобный сосуд обнаружен и на спинной стороне[2].
 Анатомия Pikaia gracilens на основе реконструкций С. Конвея Морриса и Ж.-Б. Карона (2012)
Анатомия Pikaia gracilens на основе реконструкций С. Конвея Морриса и Ж.-Б. Карона (2012)Вероятно, животное могло активно плавать, волнообразно изгибая тело, в первую очередь — его уплощённую заднюю часть[3].
Скорее всего, пикайя была фильтратором и процеживала воду со взвесью через поры своей глотки[3].
Кокако — Википедия
Материал из Википедии — свободной энциклопедии
Кокако[2][3][4][5], или гуйя-органист[3], или новозеландский скворец[3][4] (лат. Callaeas cinereus) — птица из семейства Гуйи (Callaeidae).
Длина тела около 38 см. Масса около 230 грамм. Окраска от пепельно-серой до чёрной, хвост с белой каймой.
Клюв чёрного цвета, у самца относительно короткий, у самки — более длинный, несколько изогнут книзу.
Выделяют два островных подвида[6][7]: Callaeas cinereus wilsoni с Северного острова и Callaeas cinereus cinereus (номинативный), обитавший некогда на Южном острове и острове Стьюарт, но в настоящее время считающийся уже вымершим, поскольку с 1967 года не было зафиксировано ни одного подтверждённого случая встречи этих птиц[1]. У представителей подвида Callaeas cinereus wilsoni «сережки» по бокам клюва синего цвета, а у птиц подвида Callaeas cinereus cinereus они были оранжевые. С 2010 года эти две формы орнитологами считаются отдельными видами[1][7].
Эндемик островов Новой Зеландии.
Питаются насекомыми, пауками, червями, моллюсками, ягодами, часто кормятся на земле.
Птицы гнездятся в нижних частях кроны деревьев. В кладке обычно 2—3 яйца, из которых выживает только 1—2 птенца.
16 января 2007 года южный подвид был признан вымершим[8].
На острове Стьюарт последние птицы этого подвида были отмечены в начале 1950-х годов, а на острове Южный, а также в некоторых районах западного Отаго и Нельсон — в 1960 году. Позднее появилась информация, что южноостровной подвид был встречен в 1967 году в нотофагусовых лесах, но поиски в данной местности, организованные в 1981 году не подтвердили данный факт.
Вероятно на островах Южный и Стьюарт данный подвид птиц был истреблен интродуцированными хищниками.
На островах Северный и Грейт-Барриер, по подсчетам на сентябрь 2004 года сохранилось примерно 400 пар северного подвида.
Кокако изображена на обратной стороне денежной купюры достоинством 50 новозеландских долларов.
-
Кокако двух подвидов
-


- ↑ 1 2 3 Gill B. J., Bell B. D., Chambers G. K., Medway D. G., Palma R. L., Scofield R. P., Tennyson A. J. D., Worthy T. H. 2010. Checklist of the Birds of New Zealand, Norfolk and Macquarie Islands, and the Ross Dependency, Antarctica (By the Checklist Committee Ornithological Society of New Zealand). 4-th edition. Te Papa Press, Wellington. Pp. 280—281. 501 p. ISBN 978-1-877385-59-9
- ↑ Гладков Н. А., Иноземцев А. А., Михеев А. В., Дроздов Н. Н., Ильичев В. Д., Константинов В. М., Курочкин Е. Н., Потапов Р. Л. Подотряд Певчие (Oscines) // Жизнь животных. Том 6. Птицы / под ред. В. Д. Ильичева, А. В. Михеева, гл. ред. В. Е. Соколов. — 2-е изд. — М.: Просвещение, 1986. — С. 479. — 527 с.
- ↑ 1 2 3 Галушин В. М., Дроздов Н. Н., Ильичев В. Д. и др. Фауна Мира: Птицы: Справочник / под ред. В. Д. Ильичева. — М.: Агропромиздат, 1991. — С. 290. — 311 с. — ISBN 5-10-001229-3
- ↑ 1 2 Бёме Р. Л., Флинт В. Е. Пятиязычный словарь названий животных. Птицы. Латинский, русский, английский, немецкий, французский / Под общ. ред. акад. В. Е. Соколова. — М.: Рус. яз., «РУССО», 1994. — С. 462. — 2030 экз. — ISBN 5-200-00643-0.
- ↑ Фишер Д., Саймон Н., Винсент Д. Красная книга. Дикая природа в опасности / пер. с англ., под ред. А. Г. Банникова. — М.: Прогресс, 1976. — С. 362. — 478 с.
- ↑ Paweł Mielczarek, Włodzimierz Cichocki. Polskie nazewnictwo ptaków świata. «Notatki Ornitologiczne». Tom 40. Zeszyt specjalny, 1999. ISSN 0550-0842
- ↑ 1 2 Семейство Callaeidae в Списке птиц мира Международного союза орнитологов (IOC World Bird List version 6.4)
- ↑ DoC declares South Island kokako ‘extinct’ — 16 Jan 2007 — NZ Herald: New Zealand National news
Огненногрудая петроика — Википедия
Огненногрудая петроика[1] (лат. Petroica phoenicea) — небольшая воробьиная птица родом из Австралии. Типичный обитатель районов с умеренным климатом в юго-восточной Австралии, а также острова Тасмания. Огненногрудую петроику, как и двух других представителей рода Petroica, Petroica boodang (англ.)русск. и краснолобую петроику, иногда ошибочно называют красногрудой петроикой. Длина тела от 12 до 14 см, характерны тёмно-карие глаза и тонкий чёрный клюв.
Как и у многих ярких представителей семейства австралийских зарянок, у огненногрудой петроики развит половой диморфизм. Самец имеет яркую оранжево-красную грудку и горло, а также белое пятно на лбу, в то время как самка полностью серо-коричневого цвета. Спина самца серебристо-серого цвета с белыми полосами, а хвост — чёрный с белыми кромками. Песни огненногрудой петроики описываются как самые мелодичные в рамках рода Petroica.
Происхождение огненногрудой петроики, как и австралийских зарянок, остаётся неясным, так как они не являются родственниками зарянок и странствующего дрозда. Возможно, австралийские зарянки — раннее ответвление группы Певчие воробьиные. Огненногрудая петроика преимущественно насекомоядная птица и охотится на верхушках деревьев или на земле. Огненногрудая петроика является территориальной птицей и обозначает свои владения с помощью песни или демонстрации оперения. Согласно данным международной организации BirdLife International, за последние 25 лет численность огненногрудой петроики значительно уменьшилась, и в данный момент птица находится в списке видов, близких к уязвимому положению.
Огненногрудая петроика впервые была описана в 1830 году французскими натуралистами Жаном-Рене-Констаном Куа и Жозефом-Полем Гемаром, получив название Muscicapa chrysoptera[2]. Видовое название chrysoptera происходит от др.-греч. χρυσός «золото» и πτερόν «перо»[3].
В своём описании Джон Гульд в 1837 году назвал огненногрудую петроику Petroica phoenicea, поместив её в род петроик. Именно под этим названием птица чаще всего описывалась в более поздней литературе. Название Куа и Гемара же почти не использовалось как действительное и было признано «забытым» (nomen oblitum)[4]. Название рода происходит от др.-греч. πέτρος «камень» и др.-греч. οἶκος «дом», что относится к привычке птицы сидеть на камнях[5].
Этот вид является одним из пяти представителей рода Petroica с красной или розовой грудью, которые в разговорной речи иногда именуются «Red robins» (красными робинами или красными зарянками) в противоположность «Yellow robins» (жёлтым робинам или зарянкам) — зарянковым мухоловкам[6]. Вопреки этому названию, этот вид не является близким родственником настоящей зарянки (как и странствующий дрозд, в народе называемый «американской зарянкой»)[6]. До выделения семейств Petroicidae и Eopsaltridae австралийские зарянки относились к семейству мухоловковых, а свистуны — к семейству австралийских свистунов[7][6]. Исследования Сибли (англ.)русск. и Алквиста с помощью гибридизации ДНК показали, что австралийские зарянки относятся к надсемейству Corvida, объединяющему множество тропических и австралийских воробьиных птиц, включая радужных птиц, малюровых, медососовых и врановых[8]. Тем не менее, последующие молекулярные исследования в рамках подотряда певчих воробьиных показали, что австралийские зарянки относятся к раннему ответвлению Passerida[9].
В данный момент какие-либо подвиды огненногрудой петроики не выделяются, хотя птица весьма вариабельна по окраске[10]. Взрослые самцы птиц, размножающихся на материке, описываются со светлыми брюшками и спинками, в отличие от тасманийских сородичей, однако эти различия могут быть лишь следствием старого оперения[11]. Миграция птиц через Бассов пролив только усиливает проблему выделения подвидов: материковые и тасманийские сородичи имеют те же размеры[11]. По наблюдениям орнитологов Ричарда Шодда и Яна Мейсона (англ.)русск., сложность выделения внутривидовых таксонов является следствием плохого качества музейных экспонатов и миграции[12].

Огненногрудая петроика имеет длину 12—14 см, являясь крупнейшей из австралийских зарянок[10]. Она имеет более стройное телосложение, чем другие представители рода петроик, длинные крылья и шею, а также маленькую голову[10]. Самцов легко отличить по яркому оранжево-красному оперению горла, груди и живота. Макушка, затылок, кроющие перья (англ.)русск., затылочная и боковая части шеи — тёмно-серые, а уздечка и подклювье — серо-чёрные[13]. Серые перья на макушке могут переходить в тускло-оранжевые. Остальные части тела, включая крылья, спину и хвост, — тёмно-серые[13]. Пятно над клювом, полосы на крыльях и наружные хвостовые края — белые. Перья брюшка, крыльев и клоаки белого цвета с серо-чёрной основой. Самка полностью бледно-коричневая со светло-жёлтой нижней частью тела[14]. Её туловище, бока и клоака — белые[14]. Как и у самцов, перья макушки у самок могут быть тускло-оранжевыми. Подобная особенность также может встречаться и на груди[14]. На крыльях и над клювом находятся небольшие тёмные полосы[14]. Клюв, ноги, стопы и когти чёрные, а глаза тёмно-карие[15]. В сентябре 1950 года в восточной Тасмании близ города Суонси в небольшой стае огненногрудых петроик наблюдалась особь с полностью лимонно-жёлтой грудью и оперением, отличным от остальных самок[16].
Птенцы имеют тёмно-серый или коричневый пух[14], кремовый, переходящий в серый клюв, светлый зев и оранжевую глотку[15]. Оперение молодых особей после первой линьки очень похоже на взрослое, однако голова и верхняя часть тела слегка тёмные и полосатые[14]. После второй линьки молодые особи впервые приобретают оперение, очень схожее с оперением взрослой самки[11]. В это время грудка самцов начинает покрываться оранжевыми перьями[11]. В возрасте двух лет птицы линяют в третий раз, при этом одни особи похожи на взрослых самцов, а другие сохраняют своё старое коричневое оперение[11]. Определение возраста и пола особей, имеющих коричневое оперение, является довольно затруднительной задачей[11]. Информация о точных сроках линьки отсутствует, однако замена первичного перьевого покрова происходит с декабря по февраль[15].
Цвет не является надёжным индикатором определения вида, поскольку некоторые представители Petroica boodang (англ.)русск. имеют оранжевую окраску, а самцы Petroica boodang и краснолобых петроик — красную грудь и чёрные глотки[17]. У огненногрудой петроики оперение груди простирается по бокам и заканчивается до основания клюва[17]. Огненногрудая петроика слегка более худая и имеет более маленькую голову, чем Petroica boodang, и намного крупнее, чем краснолобая петроика[17]. Самок данных видов также довольно трудно определить[11]. Самки краснолобых, розовых и малиновогрудых петроик значительно меньше, чем самый маленький представитель огенногрудой петроики с длиной крыла менее 7 см[11]. Самка Petroica boodang имеет более выраженный красный румянец на груди, а на макушке — заметное белое пятно[11].
Вокализация огненногрудой петроики состоит из громких и тихих звуков; первые можно услышать на расстоянии 150 м, а вторые — 30 м[18]. 90 % громких песен можно услышать весной, летом и осенью, в то время как с мая по июль они составляют менее 50 %[18]. Самцы в это время поют редко: они используют вокализацию только для защиты своей территории[18]. Песня огненногрудых петроик более разнообразная и сложная, чем у Petroica boodang, и является «самой мелодичной среди австралийских зарянок»[19]. Последовательность обычно состоит из трёх затухающих нот, а песня по звучанию схожа с английскими фразами: «You may come, if you will, to the sea» (рус. «вы можете, если хотите, пойти к морю») или «You are not a pretty little bird like me» (рус. «вы не такая хорошая маленькая птичка, как я»)[19]. И самцы, и самки поют эту песню, садясь на пень или забор[19]. Эта громкая песня используется либо для привлечения внимания потенциального партнёра, либо для кормёжки самки или молодняка[20]. Приглушённый зов, описанный как чередование звуков «тлип», «тёрп» или «пип», используется для связи с гнездом[20]. При приближении неприятеля к гнезду самка издаёт шипящий звук[19], а самец — хриплый голос[21].
Огненногрудая петроика встречается в умеренных регионах юго-восточной Австралии и по всей Тасмании, однако на юго-западе и западе острова она менее распространена. В штате Виктории птица обычно встречается на плоскогорьях, реже на возвышенностях[22]. Её ареал простирается от расположенных на юго-востоке Южной Австралии Аделаидских равнин и низин вокруг устья реки Муррей, далее пересекая штат Виктория, до юго-западных склонов (англ.)русск. Большого Водораздельного хребта и южных районов Нового Южного Уэльса. Севернее она встречается вдоль Большого Водораздельного хребта и его западных склонов. Несколько раз её наблюдали на юго-востоке Квинсленда[23]. В пределах своего ареала огненногрудая петроика в основном перелётная птица, перемещающаяся зимой с высокогорных и субальпийских регионов в низины, хотя обычный ареал и ареал размножения пересекаются друг с другом. Существует ряд доказательств, что самцы улетают на несколько дней раньше самок. Неясно, какая часть птиц с острова Тасмания пересекает Бассов пролив на зимовку в Виктории[24]. Птицы, которые остаются в Тасмании, улетают с мест размножения и соединяются в стаи численностью до четырнадцати птиц. Они покидают указанные области к августу, а молодые птицы улетают пораньше[25]. Полевые работы в пригороде Мельбурна Лэнгуоррин (англ.)русск. показали, что климат не влияет на большую численность огненногрудой петроики[26]. Международная организация BirdLife International в 2004 году переместила огненногрудую петроику из списка видов под наименьшей угрозой в перечень видов, близких к уязвимому положению, так как за последние 25 лет её численность сильно сократилась[27]. Правительство Австралии поместило птицу в список видов под наименьшей угрозой, хотя на границах её типичного ареала отмечалось падение численности[28]. Её популяция также поредела в Южной Австралии[29] и Виктории, хотя на самом деле это не так[28]. Они часто встречаются на высокогорьях в районе Большой Водораздельного хребта, особенно в разбросанных белых эвкалиптовых лесах и аналогичных местах обитания, а в течение периода размножения летом стабильно наблюдаются в районе Македонской горы к северо-западу от Мельбурна[28].
Весной и летом птицы чаще всего встречаются во влажных плоскогорных или высокогорных эвкалиптовых лесах на высоте до 1800 м. Они, как правило, предпочитают территории с большими полянами и небольшими подлесками. В частности, птица любит селиться в высокогорных лесах с преобладанием таких деревьев, как эвкалипт малоцветковый, эвкалипт царственный, эвкалипт прутовидный, эвкалипт косой, Eucalyptus delegatensis, эвкалипт скученный, эвкалипт Дальримпля, эвкалипт равновысокий, эвкалипт лучистый и эвкалипт миндалелистный (англ.)русск.. Иногда встречаются в умеренно влажных лесах. Осенью и зимой птицы летят на более открытые пространства, такие как поля и редколесья, около рек которых растут эвкалипт камальдульский, эвкалипт Блэкли, эвкалипт медопахнущий, эвкалипт мелкоплодный (англ.)русск. и эвкалипт железнодревесный на небольших высотах[22].
Огненногрудые петроики часто изобилуют в районах, в которых были лесные пожары, но как только появляются подлески, они улетают[22]. Они также могут прилетать на лесозаготовки или вырубленные леса[23]. Тем не менее, полевые работы в государственном лесопарке Була-Була центрального Гиппсленда показали, что птицы не встречались в районах, где была вырубка леса[30].
Огненногрудая петроика размножается в основном в районе Большого Водораздельного хребта, Тасманийского нагорья и островов в Бассовом проливе[31]. С приходом прохладной осенней погоды большинство птиц рассеивается на тёплых низменностях, а часть — в восточных районах Южной Австралии, южного Квинсленда или (в случае тасманийских птиц) Виктории, пересекая Бассов пролив. Птицы, гнездящиеся в более тёплых климатических зонах на севере Голубых гор в штате Новый Южный Уэльс, как правило, остаются на своих территориях круглый год. Вне сезона размножения птицы могут собираться в большие стаи, хотя в течение года очень часто они бывают одни или в парах. Последних чаще всего можно наблюдать во время сезона размножения[32].
Когда птица сидит на ветке или кормится на земле, она удерживает своё тело в вертикальном положении, под углом 45° и ниже, а крылья опускаются ниже хвоста. Кажется, птица создаёт впечатление нервозности и раздражения, попеременно хлопая своими крыльями, когда успокаивается[22]. Полёт птицы быстрый с заметным волнообразным характером[22].
Огненногрудая петроика — территориальная птица, защищающая свою территорию от других особей своего вида, а также от Petroica boodang (англ.)русск., которая соседствует с нею. В Ниммитабеле (англ.)русск. в районе Нового Южного Уэльса перелётные птицы вторгались и увеличивали свои территории посреди существующих владений Petroica boodang. Однажды поселившись, ни один из видов не доминировал над другим, поэтому сохранялись стабильные границы ареалов[33]. Огненногрудая петроика демонстрирует ряд черт агонистического поведения (англ.)русск., в том числе показ набухшей груди и белого пятна, подчёркивая свою белую макушку, отметины на крыльях или наружные перья на хвосте. Для защиты территорий птицы могут налетать на врагов или петь[33].
Питание[править | править код]
Как и все австралийские зарянки, огненногрудая петроика является скрытным и внезапным охотником, который питается, главным образом, насекомыми. Чтобы исследовать как можно больше корма на лесной подстилке, она часто возвращается к излюбленным низким стволам деревьев, выпрямляется и становится неподвижной. Зимой, находясь на более открытых пространствах, когда птицы довольно часто ищут еду на земле, их обычно можно видеть парами (в течение весеннего и летнего периодов размножения) или свободными группами[34]. Результаты полевых работ в районе Нового Южного Уэльса не выявили значительных различий в кормовом поведении самцов и самок[35]. Петроик также видели на свежевспаханных полях, где они охотились на насекомых, находившихся в канавках[19]. В Дениликуине наблюдалась одна огненногрудая петроика, которая во время охоты на почвенных насекомых неоднократно шуршала лапкой по земле, чтобы их вспугнуть. Как только птица замечала любое движение, она сразу же хватала свою добычу. Подобное поведение наблюдается у песочников[36].
Доля летающих насекомых в рационе огненногрудой петроики больше, чем у Petroica boodang (англ.)русск.. Биолог Дуглас Робинсон предположил, что недостаток летающих насекомых зимой является причиной миграции птиц[37]. Огненногрудые петроики наблюдались в смешанных стаях с другими мелкими насекомоядными воробьиными птицами, такими как Petroica boodang, капюшонная петроика, белолая чекановая трясогузка и степной конёк[10].
Среди насекомых потребляется много жуков, ос и муравьёв, мух (слепни и ктыри) и гусениц. Также птица питается пауками, многоножками и дождевыми червями[32]. Огненногрудая петроика в целом питается мелкими животными, а более крупную добычу перед потреблением неоднократно разбивает о твёрдую поверхность. Крупная добыча составляет только 0,5 % рациона птицы. В зависимости от пика сезонной активности осенью птица потребляет 1,8 % животных второй группы, а зимой этот показатель составляется 0,2 %[37].
Ухаживание и размножение[править | править код]
Птицы ведут преимущественно моногамный образ жизни, хотя и отмечаются случаи распада пар. Пары сохраняются на всю жизнь до тех пор, пока не погибает один из её представителей[38]. Когда самка кормится, самец либо садится рядом с ней, а затем улетает, либо летит впереди неё. Ухаживающие самцы также бегают взад и вперёд перед самкой, пряча в перья согнутые крылья и голову. В обоих случаях самец пытается преследовать самку[39].
Период размножения длится с августа по январь, во время которого бывает один или два выводка[31]. Самец ищет для самки подходящие места гнездования, прочёсывая все возможные области. В отличие от других петроик, самка иногда сама участвует в выборе гнезда. Для того, чтобы найти подходящее место гнездования, пара тратит от одного до пяти дней. Самка строит гнездо одна[21]. Эти птицы предпочитают гнездиться на эвкалиптах, но на горе Веллингтон в Тасмании их наблюдали гнездящимися на лучистой сосне. Огненногрудая петроика более непостоянна в своём выборе мест гнездования, чем другие петроики. Наблюдались случаи, когда она гнездилась в сараях[40].
Гнездо петроики представляет собой аккуратную глубокую чашу, сделанную из мягкой сухой травы, мха и коры. Для удерживания или наполнения гнезда, находящегося в развилке дерева или на скале на высоте нескольких метров от земли, используются паутина, перья и мех. Кладка, как правило, насчитывает три или четыре мутно-белых яйца, которые высиживаются несколько дней подряд[41]. Яйца с синеватым, сероватым или коричневатым оттенками и тёмно-серо-коричневыми пятнами, размером 18 на 14 мм[31]. Полевые работы на открытых эвкалиптовых лесах в Ниммитабеле показали, что огненногрудая петроика и Petroica boodang (англ.)русск. выбирают разные места гнездования: первые чаще всего селятся в дуплах деревьев и в расщелинах коры эвкалипта прутовидного на высоте около 4 м, а вторые — в развилках или на ветвях эвкалипта малоцветкового на высоте около 7 м[21]. Огненногрудые петроики, которые мигрировали в места гнездования, были более успешными в выращивании потомства, в то время как Petroica boodang были успешными в освоении территорий, бедных ресурсами[21].
Инкубационный период длится в среднем около 17 дней[21]. Как и у всех воробьиных птиц, птенцы рождаются слепыми и лысыми и начинают покрываться пухом на второй день[13]. Глаза открываются на шестой день, а на девятый или десятый птенцы начинают обрастать первичными маховыми перьями[13]. В течение первых трёх дней после вылупления мать начинает кормить птенцов, а с 4 дня — отец, сменяя друг друга в течение семи дней[42]. В пищевом рационе молодых птиц преобладают мухи, бабочки, мотыльки, гусеницы и жуки. В рационе птенцов огненногрудой петроики присутствует большая доля летающих насекомых, чем у Petroica boodang[21]. Орнитологи полагают, что это связано с более поздним началом периода размножения[21]. Оба родителя участвуют в удалении фекалий из гнезда[43]. Родители ухаживают за птенцами в течение пяти недель, после чего покидают гнездо[13].
Основными паразитами огненногрудой петроики являются веерохвостая щетинистая кукушка и бледная кукушка[13]. Самки кукушки откладывают яйца в гнёзда, которые затем высиживают петроики как свои собственные. Другими паразитами гнёзд являются серогрудая сорокопутовая мухоловка, пестрохвостая ворона-флейтист и ложные кобры Гюнтера[21].
- ↑ Бёме Р. Л., Флинт В. Е. Пятиязычный словарь названий животных. Птицы. Латинский, русский, английский, немецкий, французский / Под общ. ред. акад. В. Е. Соколова. — М.: Рус. яз., «РУССО», 1994. — С. 355. — 2030 экз. — ISBN 5-200-00643-0.
- ↑ Quoy, Jean René Constant; Gaimard, Joseph Paul in Dumont-d’Urville, J. (1830). Voyage de découvertes de l’Astrolabe exécuté par ordre du Roi, pendant les anneés 1826-1827-1828-1829, sous le commandement de M.J. Dumont-d’Urville. Zoologie. Paris: J. Tastu Vol. 1
- ↑ Liddell, Henry George; Scott, Robert. A Greek-English Lexicon (англ.)русск. (Abridged Edition) (англ.). — Oxford, United Kingdom: Oxford University Press, 1980. — P. 615, 791. — ISBN 0-19-910207-4.
- ↑ Department of the Environment, Water, Heritage and the Arts. Species Petroica (Littlera) phoenicea Gould, 1837 (неопр.) (недоступная ссылка). Australian Biological Resources Study: Australian Faunal Directory. Australian Government (9 октября 2008). Дата обращения 26 августа 2010. Архивировано 30 апреля 2013 года.
- ↑ Boles, 1988, p. 66.
- ↑ 1 2 3 Boles, 1988, p. xv.
- ↑ Boles, 1988, p. 35.
- ↑ Sibley, Charles G.; Ahlquist, Jon E. Phylogeny and Classification of Birds: A Study in Molecular Evolution (англ.). — New Haven, Connecticut: Yale University Press, 1990. — P. 603, 610—627. — ISBN 0-300-04085-7.
- ↑ Barker, F. Keith; Cibois, Alice; Schikler, Peter A.; Feinstein, Julie; Cracraft, Joel. Phylogeny and diversification of the largest avian radiation (англ.) // Proceedings of the National Academy of Sciences : journal. — 2004. — Vol. 101, no. 30. — P. 11040—11045. — DOI:10.1073/pnas.0401892101. — PMID 15263073.
- ↑ 1 2 3 4 Higgins et al., 2002, p. 666.
- ↑ 1 2 3 4 5 6 7 8 9 Higgins et al., 2002, p. 681.
- ↑ Schodde, Richard; Mason, Ian J. (англ.)русск.. The Directory of Australian Birds: Passerines. A taxonomic and zoogeographic atlas of the biodiversity of birds of Australia and its territories (англ.). — Melbourne: CSIRO Publishing (англ.)русск., 1999. — P. 273—275. — ISBN 0-643-06456-7.
- ↑ 1 2 3 4 5 6 Higgins et al., 2002, p. 678.
- ↑ 1 2 3 4 5 6 Higgins et al., 2002, p. 679.
- ↑ 1 2 3 Higgins et al., 2002, p. 680.
- ↑ Wall, L. E. Xanthochroism in scarlet robin, Petroica multicolor, and flame robin, P. phoenicea (англ.) // Emu. — Taylor & Francis, 1967. — Vol. 66, no. 3. — P. 297.
- ↑ 1 2 3 Simpson, Ken; Day, Nicolas; Trusler, Peter. Field Guide to the Birds of Australia (неопр.). — Ringwood, Victoria: Viking O’Neil, 1993. — С. 174. — ISBN 0-670-90478-3.
- ↑ 1 2 3 Higgins et al., 2002, p. 675.
- ↑ 1 2 3 4 5 North, Alfred J. Nests and eggs of birds found breeding in Australia and Tasmania (англ.). — 2nd. — Sydney: Australian Museum/F.W. White, 1903. — P. 165—167.
- ↑ 1 2 Higgins et al., 2002, p. 676.
- ↑ 1 2 3 4 5 6 7 8 Robinson, Doug. The Nesting Ecology of Sympatric Scarlet Robin Petroica multicolor and Flame Robin Petroica phoenicea Populations in Open Eucalypt Forest (англ.) // Emu : journal. — Taylor & Francis, 1990. — Vol. 90, no. 1. — P. 40—52. — DOI:10.1071/MU9900040.
- ↑ 1 2 3 4 5 Higgins et al., 2002, p. 667.
- ↑ 1 2 Higgins et al., 2002, p. 668.
- ↑ Higgins et al., 2002, p. 669.
- ↑ Thomas, D. G. Composition of Flame Robin Flocks Wintering in Tasmania (англ.) // Emu : journal. — Taylor & Francis, 1969. — Vol. 69, no. 4. — P. 240—241. — DOI:10.1071/MU969235d.
- ↑ Chambers, Lynda E. Altered timing of avian movements in a peri-urban environment and its relationship to climate (англ.) // Emu : journal. — Taylor & Francis, 2010. — Vol. 110, no. 1. — P. 48—53. — DOI:10.1071/MU09023.
- ↑ Petroica phoenicea (англ.). The IUCN Red List of Threatened Species. Дата обращения 26 ноября 2013.
- ↑ 1 2 3 Garnett, Stephen T. Taxon summary: Flame Robin (неопр.). Environment Australia. Department of the Environment, Water, Heritage and the Arts, Australian Government (2000). Дата обращения 9 августа 2010. Архивировано 1 июля 2013 года.
- ↑ Paton, D. C.; Carpenter, G.; Sinclair, R. G. A second bird atlas of the Adelaide region. Part 1: changes in the distribution of birds: 1974–75 vs 1984–85 (англ.) // South Australian Ornithologist (англ.)русск. : journal. — South Australian Ornithological Association (англ.)русск., 1994. — Vol. 31. — P. 151—193.
- ↑ Loyn, Richard H. Bird populations in a mixed Eucalypt forest used for production of wood in Gippsland, Victoria (англ.) // Emu : journal. — Taylor & Francis, 1980. — Vol. 80, no. 3. — P. 146—156.
- ↑ 1 2 3 Beruldsen, Gordon. Australian Birds: Their Nests and Eggs (неопр.). — Kenmore Hills, Queensland: self, 2003. — С. 341. — ISBN 0-646-42798-9.
- ↑ 1 2 Higgins et al., 2002, p. 672.
- ↑ 1 2 Robinson, Doug. Interspecific Aggression and Territorial Behavior Between Scarlet Robin Petroica multicolor and Flame Robin P. phoenicea (англ.) // Emu : journal. — Taylor & Francis, 1989. — Vol. 89, no. 2. — P. 93—101. — DOI:10.1071/MU9890093.
- ↑ Higgins et al., 2002, p. 671.
- ↑ Recher, Harry F.; Holmes, Richard T. The Foraging Ecology of Birds of Eucalypt Forest and Woodland. I. Differences Between Males and Females (англ.) // Emu : journal. — Taylor & Francis, 2000. — Vol. 100, no. 3. — P. 205—215. — DOI:10.1071/MU9904.
- ↑ Hobbs, John N. Flame Robin’s ‘Foot Pattering’ Feeding Habit (англ.) // Emu. — Taylor & Francis, 1954. — Vol. 54, no. 4. — P. 278—279. — DOI:10.1071/MU954278b.
- ↑ 1 2 Robinson, Doug. Why do Flame Robins Petroica phoenicea migrate? A comparison between the social and feeding ecologies of the Flame Robin and Scarlet Robin P. multicolor (англ.) // Corella : journal. — 1992. — Vol. 16. — P. 1—14.
- ↑ Robinson, Doug. The social organization of the Scarlet Robin Petroica multicolor and Flame Robin P. phoenicea in southeastern Australia: a comparison between sedentary and migratory flycatchers (англ.) // Ibis (англ.)русск. : journal. — Wiley-Blackwell (англ.)русск., 1990. — Vol. 132, no. 1. — P. 78—94. — DOI:10.1111/j.1474-919X.1990.tb
Какапо — Википедия
Кака́по[1], или сови́ный попуга́й[1] (лат. Strigops habroptila) — ночная нелетающая птица из семейства Nestoridae, являющаяся эндемиком Новой Зеландии[2]. Возможно, является одним из наиболее древних ныне живущих видов птиц[3].
 Иллюстрация какапо из книги Вальтера Лаври Буллера «Птицы Новой Зеландии» опубликованной в 1873 году[4].
Звук «бум», издаваемый какапо
Иллюстрация какапо из книги Вальтера Лаври Буллера «Птицы Новой Зеландии» опубликованной в 1873 году[4].
Звук «бум», издаваемый какапоОперение жёлто-зелёного цвета с чёрными крапинками, имеет характерный чувствительный лицевой диск, вибриссообразные перья, огромный серый клюв, короткие ноги, огромные ступни и маленькие крылья, а также относительно короткий хвост. Какапо утратил способность к активному полёту. Ведёт ночной образ жизни. У самцов и самок наблюдается половой диморфизм в размере тела. Воспитание молодняка протекает без вмешательства самца. Какапо является единственным видом попугаев, имеющим полигинийную систему размножения[5].
Длина тела около 60 см, вес в зрелом возрасте от 2 до 4 кг[6]. Оперение мягкое; окраска нижней стороны тела зеленовато-жёлтая, верхней — мшисто-зелёная, с чёрными полосами на спине. На других местах тела полосы различного цвета: жёлто-зелёные, тёмно-бурые и лимонно-жёлтые. Лицевые перья образуют лицевой диск, как у сов, и, возможно, выполняют локационную функцию. Голос хрипло-каркающий, переходящий в визгливые неприятные звуки. Одной из необычных характеристик какапо является сильный, но в то же время приятный запах, похожий на запах цветов и мёда или пчелиного воска.
Анатомия[править | править код]
Скелет какапо отличается от других попугаев, что связано с утратой способности к активному полёту. Какапо имеет наименьший относительный размер крыла среди всех попугаев. Его перья на крыльях короче, более округлые, менее асимметричные и имеют меньше дистальных штырей, соединяющих крылья вместе. Грудная клетка небольшая, имеет низкий рудиментарный киль и укороченную наружную часть позвоночника. Подобно другим нелетающим птицам вилочка какапо не срастается, а состоит из пары ключиц, соприкасающихся с каждым коракоидом. Угол состоящий из клювовидных отростков и грудины расширен. Таз более широкий чем у других представителей отряда. Проксимальные кости ног и крыльев несоразмерно длинные, а дистальные элементы укорочены[7].
Грудная мускулатура Какапо также изменена из-за утраты способности к полёту. Грудные и надкраевые мышцы значительно сокращены. Пропатагиальное сухожилие не имеет четко выраженного мышечного живота. Sternocoracoideus является сухожилием. Существует обширная мышца cucularis capitis clavicularis.[8]
Какапо был открыт английским орнитологом Джорджем Робертом Греем в 1845 году. Родовое название птицы происходит от древнегреческого strix (в родительном падеже strigos), что означает «сова», и ops — «лицо», затем видовое название habros — «мягкий», и ptilon — «перо»[9]. Он имеет так много характерных особенностей, что его даже определили в отдельную трибу Strigopini. Недавние филогенетические исследования подтвердили уникальность этого рода, а также то, что этот вид близок к видам кеа (Nestor notabilis) и новозеландский кака (Nestor meridionalis)[10][11][12]. Теперь они все определены в отдельное семейство в отряде попугаеобразных[13]. Ранее орнитологи предполагали, что какапо может быть родственен австралийским земляному попугаю (Pezoporus wallicus) и ночному попугаю (Geopsittacus occidentalis), но это мнение было опровергнуто последними исследованиями[14][15].
Был широко распространён в Новой Зеландии, в настоящее время сохранился лишь на юго-западе Южного острова. Обитают в лесах, в местах с высокой влажностью, поднимаясь до высоты 1500 м над уровнем моря. Слабость мышц и недоразвитие грудного киля диктуют птице невозможность полётов. Держатся в основном на земле. Единственный из попугаев, ведущий сумеречный и ночной образ жизни. Днём он скрывается в норах или расселинах скал. Ночью по протоптанным тропам выходит кормиться ягодами или соком растений (жуёт листья и побеги, не срывая их).
Единственными коренными млекопитающими являются три вида маленьких летучих мышей, один из которых вымер. Видимо, какапо, как многие другие птицы, заменяет млекопитающих, делая работу, присущую им. До людей какапо был распространён шире и на всех трёх главных островах Новой Зеландии. Они обитали в различных местностях, включая дерновину, кустарниковые заросли и береговые районы. Также они населяют леса, включая те местности, где растут ногоплодниковые растения (Podocarpaceae), например, дакридиум кипарисовый, или риму (Dacrydium cupressinum), псеудотсуга тисолистная (Prumnopitys taxifolia), ногоплодник дакридиевидный (Dacrycarpus dacrydioides) и тотара (Podocarpus totara), а также нотофагус (Nothofagus), тава (Beilschmiedia tawa) и метросидерос зонтичный (Metrosideros umbellata). На фиордах, в районах обвала и скалистых развалинах с регенерирующейся и с тяжёлой растительностью (например, винная ягода, или макомако (Aristotelia serrata), новозеландские виды рубус, или малинника (Rubus), кориария (Coriaria), хебе (Hebe) и копросмы (Coprosma)), называются «садом какапо»[16].
Какапо — в первую очередь, ночной попугай. Он делает насест под укрытием дерева или на земле на день, а с наступлением ночи начинает обходить свою территорию[2].
Хоть какапо и не умеют летать, зато они отличные альпинисты и могут вскарабкаться на вершину высокого дерева. Затем могут спускаться с ‘‘парашютом’’, то есть спускаться спрыгивая с раскрытыми крыльями. Таким образом они могут путешествовать (планировать) на 20-50 метров под углом 45°[5].
Питание[править | править код]
Клюв какапо приспособлен для измельчения пищи. По этой причине птица имеет очень маленькую глотку по сравнению с другими птицами подобного размера. В основном он травоядный, питается местными растениями, семенами, ягодами, пыльцой и даже заболонью. Исследования питания 1984 году открыли где-то 25 видов растений, которыми питается птица[2]. Какапо очень любят фрукты риму и питаются ими весь сезон года.
Диета какапо меняется каждый сезон. На протяжении всего года они питаются следующими растениями: Lycopodium ramulosum, Lycopodium fastigium, Schizaea fistulosa, Blechnum minus, Blechnum procerum, Cyathodes juniperina, Dracophyllum longifolium, Olearia colensoi и Thelymitra venosa. Отношение к некоторым видам растений и птиц бывает разным. Какапо оставляет явные следы своего пребывания на местах кормления на участке размером от 10×10 м до 50×100 м[2]. Манука (Leptospermum scoparium) и кусты Lepidothamnus intermedius имеют явные признаки излюбленного корма.
Размножение[править | править код]
Гнёздами им служат выкопанные ямки в сердцевине гнилого пня или дерева, а также расщелины скал. Случается, что в гнездовую нору ведут два входа, от которых отходят туннели, имеющие в длину несколько десятков сантиметров, а в глубине оборудуется камера. Яйца откладываются в январе-феврале. В кладке обычно 2 яйца. Самцы какапо призывают самок звуком необычайно низкого тона, типа урчания, слышного на несколько километров.
Какапо находится в категории CR (таксоны в критическом состоянии). На январь 2019 года известно 147 особей[17], все особи известны, многим из них были даны имена[18]. Близкими родственными видами совиному попугаю являются попугаи из рода Nestor[10][11][12]. Какапо стал изолированным от них, когда Новая Зеландия отделилась от Гондваны примерно 82 миллиона лет назад. А примерно 70 миллионов лет назад какапо удалился от представителей рода Nestor. Из-за отсутствия на территории Новой Зеландии млекопитающих хищников какапо потерял способность летать. Из-за колонизации островов полинезийцами и европейцами, которые завезли на остров крыс, котов и горностаев, большое количество особей совиного попугая было истреблено.
Находится на грани исчезновения. В Красной книге МСОП. Некоторые учёные считают, что в настоящее время в природе эти попугаи, возможно, вымерли.
Ещё в XIX веке стало ясно, что какапо находится на грани вымирания. В то время люди отлавливали птиц и отвозили их на остров Резольюшен. Всего туда было отправлено 300 попугаев, большая часть которых стала жертвой мелких хищников. Сведение, что какапо слышали в 1961 году на острове Северный, вероятнее всего, ошибочны. Хотя в 1960 году численность особей насчитывала почти 200 птиц.
Основная причина их исчезновения — деятельность интродуцированных грызунов и хищников (включая крыс и кошек), а также уничтожение и изменение их местообитания человеком и завезёнными на остров копытными. Почти все птицы, отловленные в 60-70 гг., оказались самцами, самок ловили лишь на острове Стьюарт. Д. Мертон объясняет преобладание самцов в природе тем, что какапо является представителем токовых птиц.
Помимо запрета добычи этих птиц и включения их в Приложение 1 СИТЕС, одним из возможных путей его сохранения является расселение какапо на острова, свободные от хищников, — своеобразные авиарии.
В 1974—1975 гг. 3-х попугаев (возможно, самцов) выпустили на остров Мауд, расположенный у северо-восточных берегов острова Южный в границах морского парка Марлоборо-Саундс; предполагалось интродуцировать какапо на Литл-Барриер.
В 1999 году насчитывалось всего 62 совиных попугая, в том числе 6 птенцов. Департамент охраны природы Новой Зеландии считает своей главной задачей спасение данного вида, поэтому птиц разводят в неволе, а затем выпускают на защищённые территории. Пар они не образуют, а вырытые птицами группы ям используются многими самцами как места демонстрации себя перед самками, посещающие эти «токовые площадки». Поэтому разведение этих птиц в неволе представляет значительные трудности, хотя вольерное содержание они переносят довольно хорошо, что было отмечено ещё в XIX веке.
В 2009 году был значительный брачный сезон у какапо, и родилось 33 птенца[19]. Количество особей достигло 100 штук. В 2016 также родилось много птенцов (32 особи)[19]. В остальные годы прироста популяции либо не было, либо был незначительный прирост 2-5 птенцов. К 2017 году на островах было известно 153 птицы, а в 2018 уже 147. По данным организации Kakapo Recovery в 2019 году ожидается сезон размножения, и организация надеется на 30-50 птенцов[17].
Вмешательство человека[править | править код]
Первым фактором снижения популяции является прибытие людей. Согласно фольклору народа маори, какапо жили во всех местностях, пока полинезийцы не прибыли в страну длинного белого облака (Aotearoa — синоним Новой Зеландии) 1000 лет назад[20]. Второй фактор — ввоз в страну инвазионных видов европейцами.
План по восстановлению численности какапо[править | править код]
| Перемещён | Численность | Умершие < 6 месяцев | Выжившие до ноября 1992 года |
|---|---|---|---|
| Остров Мауд (1974-81) | 9 (6♂, 3♀) | 3 (2♂, 1♀) | 4 (2♂, 2♀) |
| Маленький Барьерный Остров (1982) | 22 (13♂, 9♀) | 2 (1♂, 1♀) | 15-19 (10-12♂, 5-7♀) |
| Остров Кодфиш (1987-92) | 30 (20♂, 10♀) | 0 | 20-30 (13-20♂, 7-10♀) |
| Остров Мауд (1989-91) | 6 (4♂, 2♀) | 0 | 5 (3♂, 2♀) |
| Остров Мана (1992) | 2 (2♀) | 1 (1♀) | 1 (1♀) |
| Общее | 65 (43♂, 22♀) | 6 (3♂, 3♀) | 41-55 (27-36♂, 14-19♀) |
| Заметка: ♂ = самцы, ♀ = самки. | |||
Очень умные и добродушные попугаи, за доброе отношение они очень сильно привязываются к человеку, выражая ему свою любовь и привязанность, подобно собакам и кошкам, ласкающимся к своему хозяину.[источник не указан 16 дней]
Какапо, как и другие виды птиц, является важной частью истории для народа маори (коренные жители островов Новой Зеландии). Маори слагали о них легенды и использовали в фольклоре. Мясо какапо использовалось народом маори в пищу и считается деликатесом[22], поэтому они охотились на птиц, пока те были широко распространены.
- ↑ 1 2 Бёме Р. Л., Флинт В. Е. Пятиязычный словарь названий животных. Птицы. Латинский, русский, английский, немецкий, французский / Под общ. ред. акад. В. Е. Соколова. — М.: Рус. яз., «РУССО», 1994. — С. 129. — 2030 экз. — ISBN 5-200-00643-0.
- ↑ 1 2 3 4 H.A. Best. The Foods of Kakapo on Stewart Island as Determined from Their Feeding Sign (англ.) // New Zealand Ecological Society : journal. — New Zealand Journal of Ecology (англ.)русск., 1984. — Vol. 7. — P. 71—83.
- ↑ Ralph G. Powlesland, Don V. Merton, and John F. Cockrem. A parrot apart: the natural history of the kakapo (Strigops habroptila), and the context of its conservation management (англ.) // Notornis : journal. — 2006. — Vol. 53, no. 1. — P. 3—26.
- ↑ New Zealand Birds | Birds | Gallery | Kakapo, Strigops habroptilus
- ↑ 1 2 Ralph G. Powlesland, Don V. Merton, and John F. Cockrem. A parrot apart: the natural history of the kakapo (Strigops habroptila), and the context of its conservation management (англ.) // Notornis : journal. — 2006. — Vol. 53, no. 1. — P. 3—26.
- ↑ Higgins, P.J. Handbook of Australian, New Zealand and Antarctic Birds (англ.)русск.. Volume 4: Parrots to Dollarbird (англ.). — Melbourne: Oxford University Press, 1999. — ISBN 0-19-553071-3.
- ↑ a b Livezey, Bradley C. (July 1992). «Morphological corollaries and ecological implications of flightlessness in the kakapo (Psittaciformes: Strigops habroptilus)». Journal of Morphology. 213 (1): 105–145.
- ↑ a b Livezey, Bradley C. (July 1992). «Morphological corollaries and ecological implications of flightlessness in the kakapo (Psittaciformes: Strigops habroptilus)». Journal of Morphology. 213 (1): 105–145.
- ↑ Liddell, Henry George and Robert Scott (англ.)русск.. A Greek-English Lexicon (англ.)русск. (Abridged Edition) (англ.). — United Kingdom: Oxford University Press, 1980. — ISBN 0-19-910207-4.
- ↑ 1 2 Wright, T.F.; Schirtzinger E. E., Matsumoto T., Eberhard J. R., Graves G. R., Sanchez J. J., Capelli S., Muller H., Scharpegge J., Chambers G. K. & Fleischer R. C. A Multilocus Molecular Phylogeny of the Parrots (Psittaciformes): Support for a Gondwanan Origin during the Cretaceous (англ.) // Mol Biol Evol : journal. — 2008. — Vol. 25, no. 10. — P. 2141—2156. — DOI:10.1093/molbev/msn160. — PMID 18653733.
- ↑ 1 2 Grant-Mackie, E.J.; J.A. Grant-Mackie, W.M. Boon & G.K. Chambers. Evolution of New Zealand Parrots (неопр.) // NZ Science Teacher. — 2003. — Т. 103.
- ↑ 1 2 de Kloet, R.S.; de Kloet, S.R. (2005). The evolution of the spindlin gene in birds: sequence analysis of an intron of the spindlin W and Z gene reveals four major divisions of the Psittaciformes. Molecular Phylogenetics and Evolution 36: 706—721.
- ↑ Christidis L., Boles W. E. Systematics and Taxonomy of Australian Birds (англ.). — Canberra: CSIRO Publishing (англ.)русск., 2008. — P. 200. — ISBN 9780643065116.
- ↑ Schodde, R. & Mason, I.J. (1981). Nocturnal Birds of Australia. Illustrated by Jeremy Boot. Melbourne: Lansdowne Edns 136 pp. 22 pls [35-36]
- ↑ Leeton, P.R.J., Christidis, L., Westerman, M. & Boles, W.E. (1994). Molecular phylogenetic relationships of the Night Parrot (Geopsittacus occidentalis) and the Ground Parrot (Pezoporus wallicus). Auk 111: 833—843
- ↑ P. N. Johnson. Vegetation associated with kakapo (Strigops habroptilus Gray) in Sinbad Gully, Fiordland, New Zealand (англ.) // New Zealand Journal of Botany : journal. — 1975. — Vol. 14. — P. 151—159. Архивировано 11 апреля 2008 года. Архивная копия от 11 апреля 2008 на Wayback Machine
- ↑ 1 2 Kākāpō Recovery (нз. англ.) (неопр.) ?. www.doc.govt.nz. Дата обращения 8 февраля 2019.
- ↑ KAKAPO PARROTS — The 86 Names (неопр.). anotherchancetosee.com (4 августа 2006). Дата обращения 6 февраля 2007. Архивировано 18 февраля 2012 года.
- ↑ 1 2 The history of kākāpō (нз. англ.) (неопр.) ?. www.doc.govt.nz. Дата обращения 8 февраля 2019.
- ↑ Rob Tipa. Kakapo in Māori lore (англ.) // Notornis. — 2006. — Vol. 53, no. 1.
- ↑ B. D. Lloyd and R. G. Powlesland. The decline of kakapo Strigops habroptilus and attempts at conservation by translocation (англ.) // Biological Conservation : journal. — 1994. — Vol. 69, no. 1. — P. 75—85. — DOI:10.1016/0006-3207(94)90330-1.
- ↑ Rob Tipa, Short note: «Kakapo in Māori Lore», Notornis, Vol. 53, 193—194 (неопр.) (недоступная ссылка). Дата обращения 10 августа 2009. Архивировано 11 апреля 2008 года.
- Биологический энциклопедический словарь / Гл. ред. М. С. Гиляров; Редкол.: А. А. Баев, Г. Г. Винберг, Г. А. Заварзин и др. — 2-е изд., исправл. — М.: Советская энциклопедия, 1989. — 864 с. — ISBN 5-85270-002-9 (См. аннотацию.)
- Иерусалимский И. Попугаи и певчие птицы: виды, содержание, обучение. — Ростов-на-Дону: Владис, 2001. — 320 с. — ISBN 5-94194-073-4
- Остапенко В. А. Птицы в вашем доме: Справочное пособие. — М.: Арнадия, 1996. — ISBN 5-88666-011-9
- Butler, David. Quest for the kakapo (неопр.). — Auckland: Heinemann Reed (англ.)русск., 1989. — ISBN 0-7900-0065-2.
- Climo, Gideon and Ballance, Alison. Hoki: the story of a kakapo (неопр.). — Auckland: Godwit, 1997. — ISBN 1-86962-009-7.
- Jones, Jenny. The Kakapo (неопр.). — Auckland: Reed, 2003. — ISBN 1-86948-662-5.
- IUCN 2006 (Database entry includes justification for why this species is critically endangered.)
- Murray Williams and Don Merton. Saving Kakapo: an illustrated history, 2006 (англ.) // Notornis. — Vol. 53, no. 1.
- Eulenpapagei oder Kakapo (Strigops habroptilus). in: Günther Steinig (Hrsg.): Brehms Exotische Vogelwelt. Safari, Berlin ²1963, S.62-71. (Die Darstellung folgt vor allem Beobachtungen frühen Erforschern Neuseelands, wie Julius Haast, Georg Grey und Lyall)
- Jim Rearden: Die letzten Tage des Kakapo. in: Geo-Magazin. Hamburg 1978,2, S.88-102. (über die Erhaltungsbemühungen in Fiordland). ISSN 0342-8311
- Vom Leben eines totgesagten Vogels. in: Geo-Magazin. Hamburg 2006,10(Okt.), S.176-180. ISSN 0342-8311
- Eulenpapagei. Brummend balzt das letzte Männchen. in: R. L. Schreiber, A. W. Diamond, H. Stern, G. Thielcke (Hrsg.): Rettet die Vogelwelt. O. Maier, Ravensburg 1987, S.198-201. ISBN 3-473-46160-1
- Douglas Adams, Mark Carwardine: Die Letzten ihrer Art — Eine Reise zu den aussterbenden Tieren unserer Erde. Hoffmann und Campe, Hamburg 1991. ISBN 3-455-08384-6
- Don V. Merton, Rodney B. Morris, Ian A. E. Atkinson: Lek behaviour in a parrot: the Kakapo Strigops habroptilus of New Zealand. in: The Ibis. Oxford 126.1984. ISSN 0019-1019
- David Cemmick, Dick Veitch: Kakapo Country. The Story of the World’s most unusual bird. Foreword by David Bellamy. Illustrationen von D. Cemmick. Hodder&Stoughton, Auckland 1987. ISBN 0-340-41647-5
- Rod Morris, Hal Smith: Wild South. Saving New Zealand’s Endangered Birds. TVNZ and Century Hutchinson, Auckland 1988. ISBN 1-86941-043-2
- Philip Temple, Chris Gaskin: The Story of the kakapo. Parrot of the Night. Hodder&Stoughton, Auckland 1988. (Pricewinner: Children’s Picture Book of the Year Award 1990). ISBN 0-340-51967-3
- Ralph Powlesland: Kakapo Recovery Plan 1989—1994. Published by The Department of Conservation (DoC), Wellington 1989. ISBN 0-478-01114-8
- R. G. Powlesland, A. Roberts, B. D. Lloyd, D. Merton: Number, fate, and distribution of Kakapo (Strigops habroptilus) found on Stewart Island, New Zealand 1979—1992. in: New Zealand Journal of Zoology. Wellington 22.1995, 239—248. ISSN 0301-4223
- Mary Cresswell, Kakapo Management Group: KAKAPO RECOVERY PLAN 1996—2005. Threatened Species Recovery Plan No. 21. Department of Conservation (DoC), Wellington 1996. ISBN 0-478-01773-1
- Don Merton: Kakapo. in: P. J. Higgins (Hrsg.): Handbook of Australian, New Zealand and Antarctic Birds. Bd 4. RAOU. Oxford University Press, Melbourne 1999, 633—646. ISBN 0-19-553071-3
- Tim Higham: The Kakapo of Codfish Island. in: New Zealand Geographic magazine. Auckland 1992,15 (July-Sept.), 30-38. ISSN 0113-9967
- Derek Grzelewski: Kakapo. Bird on the brink. in: New Zealand Geographic Magazine. Ohakune 2002, 56 (March-April). ISSN 0113-9967
- Gerard Hutching: Back from the Brink. The Fight to Save our Endangered Birds. Penguin Books Publisher, Auckland 2004. ISBN 0-14-301948-1
- A celebration of kakapo. Special Issue of Notornis. Ornithological Society of New Zealand, Wellington 53.2006,1. ISSN 0029-4470
Picozoa — Википедия
Материал из Википедии — свободной энциклопедии
| Picozoa | |
|---|---|
| Picomonas judraskeda | |
Picozoa Seenivasan, Sausen, Medlin & Melkonian, 2013 | |
Picozoa (ранее Picobiliphyta) — тип[1] морских планктонных протистов неясного систематического положения. По состоянию на 2015 год, в типе описан один вид Picomonas judraskeda. Кроме того, к данной группе относят большое количество последовательностей генов рибосомных РНК, полученных с помощью методов метагеномики[2].
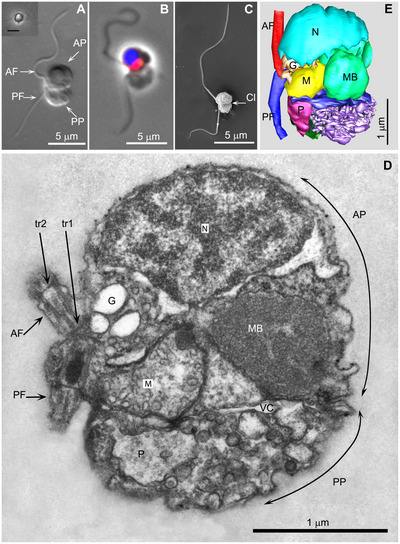
В 2007 году Фабрис Но (лат. Fabrice Not) с соавторами опубликовали статью об обнаружении ранее неизвестной клады протистов высокого ранга на основании филогенетического анализа последовательностей генов рибосомных РНК, выделенных из морских планктонных проб верхних слоев толщи воды. Изучив клетки данных организмов, выявленные с помощью методики TSA-FISH, авторы предположили наличие у них фотосинтезирующих пластид, организованных по типу хлоропластов криптофитовых водорослей и обладающих особой ДНК-содержащей органеллой — нуклеоморфом[en]. На основании этого авторы характеризовали данную группу как представителей фитопланктона и присвоили ей название Picobiliphyta[2].
Однако в дальнейшем фототрофный образ жизни данных организмов был поставлен под сомнение, так как исследования, направленные на выявление у Picobiliphyta пластид или фрагментов ДНК, содержащейся в данных органеллах, не принесли результатов. Кроме того, представители данного таксона были обнаружены в морских пробах, отобранных на глубине более 3000 метров, где отсутствие света делает невозможным фотосинтез[3].
Окончательным доказательством того, что Picobiliphyta являются гетеротрофами, стало описание в 2013 году первого представителя таксона — Picomonas judraskeda. Авторы продемонстрировали, что данный организм не способен к фотосинтезу и питается, поглощая крохотные (менее 150 нанометров) органические частицы из толщи воды. На основании этих данных авторы опубликовали формальное таксономическое описание группы, присвоив ей ранг типа и переименовав из Picobiliphyta в Picozoa, в соответствии с образом жизни данных протистов[1].
Размер клеток P. judraskeda, также как и других не описанных представителей Picozoa, выделенных из планктонных проб, составляет около 3 микрометров. Строение данных организмов уникально и не имеет аналогов среди прочих эукариот. Клетка P. judraskeda разделена глубокой щелевидной впадиной на две части, сильно отличающихся морфологически и функционально. Одна часть содержит основной набор клеточных органелл: ядро, одиночную митохондрию с трубчатыми кристами, 2 крупных микротельца (мембранных органелл, вероятно связанных с метаболизмом липидов), аппарат Гольджи, эндоплазматический ретикулум, жгутиковый аппарат (два жгутика неравного размера располагаются на боковой стороне клетки). Вторая часть специализируется на питании и содержит «клеточный рот» (цитостом), набор пищеварительных везикул и вакуолей. Внутри клетки эти две части разделены крупной вокуолярной цистерной с неизвестной функцией, возможно, являющейся барьером для попадания в клетку вирусов вместе с пищей[1][3].
Видеозапись движения клетки P. judraskedaДля P. judraskeda описан уникальный тип движения в толще воды — так называемый цикл «jump, drag, and skedaddle» (дословный перевод — прыжок, волочение и улепетывание). После длительного периода покоя клетка делает резкий рывок на небольшое расстояние и начинает медленное плавное движение в противоположном направлении. Эта часть цикла может быть повторена несколько раз, после чего Picomonas чрезвычайно быстро уплывает на большое расстояние от его изначального положения, и цикл начинается снова[1].
Вероятное систематическое положение[править | править код]
Систематическое положение Picozoa относительно других групп эукариот доподлинно не известно. Клетки описанного вида имеют уникальное строение и не похожи на других протистов. Молекулярно-филогенетические исследования данной группы также не дают однозначной информации. Из-за того что до 2013 года группу считали представителями фитопланктона, Picozoa в первую очередь сравнивали с представителями других таксонов водорослей. По состоянию на 2014 год были предложены следующие варианты систематического положения данной группы[3]:
|
|
|
|
- ↑ 1 2 3 4 5 Seenivasan, R.; Sausen, N.; Medlin, L. K.; Melkonian, M. Picomonas judraskeda Gen. Et Sp. Nov.: The First Identified Member of the Picozoa Phylum Nov., a Widespread Group of Picoeukaryotes, Formerly Known as ‘Picobiliphytes’ (англ.) // PLoS ONE. — 2013. — Vol. 8. — P. e59565. — DOI:10.1371/journal.pone.0059565.
- ↑ 1 2 3 Not F., Valentin K., Romari K., Lovejoy C., Massana R., et al. Picobiliphytes: a marine picoplanktonic algal group with unknown affinities to other eukaryotes // Science. — 2007. — Vol. 315. — P. 253—255. — DOI:10.1126/science.1136264.
- ↑ 1 2 3 Moreira D., López-García P.,. The rise and fall of Picobiliphytes: How assumed autotrophs turned out to be heterotrophs // Bioessays. — 2014. — Vol. 36. — P. 468–474. — DOI:10.1002/bies.201300176.
- ↑ Cuvelier ML, Ortiz A, Kim E, Moehlig H, Richardson DE, et al. Widespread distribution of a unique marine protistan lineage // Environ Microbiol. — 2008. — Vol. 10. — P. 1621–1634. — DOI:10.1111/j.1462-2920.2008.01580.x.
- ↑ Yoon HS, Price DC, Stepanauskas R, Rajah VD, Sieracki ME, et al. Single-cell genomics reveals organismal interactions in uncultivated marine protists. // Science. — 2011. — Vol. 332. — P. 714-717. — DOI:10.1126/science.1203163.
- ↑ Burki F, Okamoto N, Pombert JF, Keeling PJ. The evolutionary history of haptophytes and cryptophytes: phylogenomic evidence for separate origins. // Proc Biol Sci.. — 2012. — Vol. 279. — P. 2246–2254. — DOI:10.1098/rspb.2011.2301.